La señalización autocrina es una forma de señalización celular en la que una célula secreta una hormona o mensajero químico (denominado agente autocrino) que se une a receptores autocrinos en esa misma célula, lo que provoca cambios en la misma. [ 1 ] Esto se puede contrastar con la señalización paracrina , la señalización intracrina o la señalización endocrina clásica .
Ejemplos
Un ejemplo de agente autocrino es la citocina interleucina-1 en los monocitos . Cuando la interleucina-1 se produce en respuesta a estímulos externos, puede unirse a receptores de la superficie celular en la misma célula que la produjo.
Otro ejemplo se da en los linfocitos T activados , es decir, cuando un linfocito T madura al unirse a un complejo péptido : MHC en una célula presentadora de antígeno profesional y mediante la señal coestimuladora B7 : CD28 . Tras la activación, los receptores de IL-2 de baja afinidad se reemplazan por receptores de IL-2 de alta afinidad que constan de cadenas α, β y γ. La célula libera IL-2, que se une a sus nuevos receptores de IL-2, provocando autoestimulación y, finalmente, una población monoclonal de linfocitos T. Estos linfocitos T pueden entonces realizar funciones efectoras como la activación de macrófagos , la activación de linfocitos B y la citotoxicidad mediada por células .
Cáncer
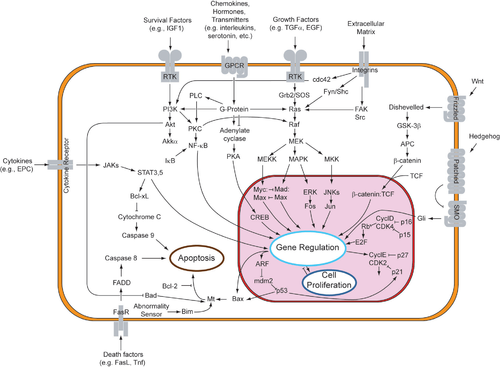
El desarrollo tumoral es un proceso complejo que requiere división, crecimiento y supervivencia celular . Una estrategia que utilizan los tumores para aumentar su crecimiento y supervivencia es la producción autocrina de factores de crecimiento y supervivencia. La señalización autocrina desempeña un papel fundamental en la activación del cáncer y también en el suministro de señales de crecimiento autosostenible a los tumores.
En la vía Wnt
Normalmente, la vía de señalización Wnt conduce a la estabilización de la β-catenina mediante la inactivación de un complejo proteico que contiene los supresores tumorales APC y Axin . Este complejo de destrucción normalmente desencadena la fosforilación de la β-catenina , induciendo su degradación. La desregulación de la vía de señalización Wnt autocrina a través de mutaciones en APC y Axin se ha relacionado con la activación de varios tipos de cáncer humano . [ 2 ] [ 3 ] Las alteraciones genéticas que conducen a la desregulación de la vía Wnt autocrina resultan en la transactivación del receptor del factor de crecimiento epidérmico (EGFR) y otras vías, lo que a su vez contribuye a la proliferación de células tumorales. En el cáncer colorrectal , por ejemplo, las mutaciones en APC, axin o β-catenina promueven la estabilización de la β-catenina y la transcripción de genes que codifican proteínas asociadas al cáncer . Además, en el cáncer de mama humano , la interferencia con la vía de señalización Wnt desregulada reduce la proliferación y la supervivencia del cáncer. Estos hallazgos sugieren que la interferencia con la señalización Wnt a nivel del ligando-receptor puede mejorar la eficacia de las terapias contra el cáncer. [ 3 ]
IL-6
La interleucina 6 (acrónimo: IL-6) es una citocina importante para muchos aspectos de la biología celular , incluyendo las respuestas inmunitarias , la supervivencia celular , la apoptosis y la proliferación . [ 4 ] Varios estudios han resaltado la importancia de la señalización autocrina de IL-6 en los cánceres de pulmón y mama. Por ejemplo, un grupo encontró una correlación positiva entre la STAT3 fosforilada en tirosina persistentemente activada (pSTAT3), presente en el 50% de los adenocarcinomas de pulmón, y la IL-6. Investigaciones posteriores revelaron que el EGFR mutante podría activar la vía oncogénica de STAT3 a través de la señalización autocrina de IL-6 regulada al alza. [ 5 ]
De manera similar, la sobreexpresión de HER2 ocurre en aproximadamente una cuarta parte de los cánceres de mama y se correlaciona con un mal pronóstico. Investigaciones recientes revelaron que la secreción de IL-6 inducida por la sobreexpresión de HER2 activó STAT3 y alteró la expresión génica, lo que resultó en un bucle autocrino de expresión de IL-6/STAT3. Tanto los modelos in vivo de cáncer de mama con sobreexpresión de HER2 en ratones como en humanos dependieron críticamente de esta vía de señalización HER2–IL-6–STAT3. [ 6 ] Otro grupo encontró que los altos niveles séricos de IL-6 se correlacionaban con un mal pronóstico en tumores de cáncer de mama. Su investigación mostró que la señalización autocrina de IL-6 indujo características malignas en mamósferas que expresan Notch-3. [ 7 ]
IL-7
Un estudio demuestra cómo la producción autocrina de la citocina IL-7 mediada por la leucemia linfoblástica aguda de células T (LLA-T) puede estar implicada en el desarrollo oncogénico de la LLA-T y ofrece nuevas perspectivas sobre la propagación de la LLA-T. [ 8 ]
VEGF
Otro agente implicado en la señalización autocrina del cáncer es el factor de crecimiento endotelial vascular (VEGF). El VEGF, producido por las células del carcinoma, actúa mediante señalización paracrina en las células endoteliales y mediante señalización autocrina en las células del carcinoma. [ 9 ] La evidencia muestra que el VEGF autocrino está implicado en dos aspectos principales del carcinoma invasivo: la supervivencia y la migración. Además, se ha demostrado que la progresión tumoral selecciona células dependientes de VEGF, lo que cuestiona la creencia de que el papel del VEGF en el cáncer se limita a la angiogénesis . En cambio, esta investigación sugiere que las terapias dirigidas al receptor de VEGF pueden afectar la supervivencia y la invasión del cáncer, así como la angiogénesis. [ 9 ] [ 10 ]
Promoción de la metástasis
La metástasis es una causa importante de muertes por cáncer, y faltan estrategias para prevenir o detener la invasión. Un estudio mostró que la señalización autocrina de PDGFR juega un papel esencial en el mantenimiento de la transición epitelio-mesenquimal (EMT) in vitro, que se sabe que se correlaciona bien con la metástasis in vivo. Los autores mostraron que el potencial metastásico de las células epiteliales mamarias oncogénicas requería un bucle de señalización autocrina PDGF/PDGFR, y que la cooperación de la señalización autocrina de PDGFR con el oncogén era necesaria para la supervivencia durante la EMT. La señalización autocrina de PDGFR también contribuye al mantenimiento de la EMT, posiblemente a través de la activación de STAT1 y otras vías distintas. Además, la expresión de PDGFRα y -β se correlacionó con el comportamiento invasivo en carcinomas mamarios humanos. [ 11 ] Esto indica las numerosas vías a través de las cuales la señalización autocrina puede regular los procesos metastásicos en un tumor.
Desarrollo de objetivos terapéuticos
El creciente conocimiento sobre el mecanismo de la señalización autocrina en la progresión del cáncer ha revelado nuevos enfoques para el tratamiento terapéutico. Por ejemplo, la señalización autocrina de Wnt podría proporcionar una nueva diana para la intervención terapéutica mediante antagonistas de Wnt u otras moléculas que interfieran con las interacciones ligando - receptor de la vía de Wnt. [ 2 ] [ 3 ] Además, la producción de VEGF-A y la activación de VEGFR-2 en la superficie de las células de cáncer de mama indican la presencia de un bucle de señalización autocrina distinto que permite a las células de cáncer de mama promover su propio crecimiento y supervivencia mediante la fosforilación y activación de VEGFR-2. Este bucle autocrino es otro ejemplo de una diana terapéutica atractiva . [ 9 ]
En los cánceres de mama con sobreexpresión de HER2, la relación de señalización HER2–IL-6–STAT3 podría ser un objetivo para desarrollar nuevas estrategias terapéuticas. [ 6 ] Los inhibidores de la quinasa HER2, como el lapatinib, también han demostrado eficacia clínica en cánceres de mama con sobreexpresión de HER2 al interrumpir un bucle autocrino mediado por la neuregulina-1 (NRG1). [ 12 ]
En el caso de la señalización de PDGFR, la sobreexpresión de un PDGFR dominante negativo o la aplicación del fármaco contra el cáncer STI571 son dos enfoques que se están explorando para interferir terapéuticamente con la metástasis en ratones. [ 11 ]
Además, se pueden desarrollar fármacos que activen la señalización autocrina en las células cancerosas, la cual de otro modo no se produciría. Por ejemplo, se ha demostrado que un mimético de molécula pequeña de Smac /Diablo, que contrarresta la inhibición de la apoptosis, potencia la apoptosis inducida por fármacos quimioterapéuticos mediante la secreción autocrina del factor de necrosis tumoral alfa (TNFα). En respuesta a la señalización autocrina de TNFα, el mimético de Smac promueve la formación de un complejo activador de la caspasa-8 dependiente de RIPK1, lo que conduce a la apoptosis. [ 13 ]
Papel en la resistencia a los medicamentos
Estudios recientes han revelado la capacidad de las células cancerosas resistentes a los fármacos para adquirir señales mitogénicas a partir de bucles autocrinos previamente ignorados, lo que provoca la recurrencia del tumor.
Por ejemplo, a pesar de la expresión generalizada de receptores del factor de crecimiento epidérmico (EGFR) y ligandos de la familia EGF en el cáncer de pulmón de células no pequeñas (CPCNP), los inhibidores de la tirosina quinasa específicos de EGFR, como el gefitinib, han mostrado un éxito terapéutico limitado. Se propone que esta resistencia se debe a que en las células de CPCNP se activan vías de señalización de crecimiento autocrino distintas del EGFR. El análisis del perfil de expresión génica reveló la prevalencia de factores de crecimiento de fibroblastos (FGF) específicos y receptores de FGF en líneas celulares de CPCNP, y se encontró que FGF2, FGF9 y sus receptores abarcan un bucle autocrino de factor de crecimiento que está activo en un subconjunto de líneas celulares de CPCNP resistentes al gefitinib. [ 14 ]
En el cáncer de mama, la adquisición de resistencia al tamoxifeno constituye otro problema terapéutico importante. Se ha demostrado que la fosforilación de STAT3 y la expresión de RANTES aumentan en respuesta al tamoxifeno en células de cáncer de mama humanas. En un estudio reciente, un grupo demostró que STAT3 y RANTES contribuyen al mantenimiento de la resistencia al fármaco mediante la regulación positiva de señales antiapoptóticas y la inhibición de la escisión de caspasas . Estos mecanismos de señalización autocrina STAT3-RANTES sugieren una estrategia novedosa para el manejo de pacientes con tumores resistentes al tamoxifeno. [ 15 ]
Véase también
- La señalización paracrina es una forma de comunicación intercelular en la que una célula produce una señal para inducir cambios en las células cercanas, alterando el comportamiento o la diferenciación de estas últimas.
- Intracrina
- Hormona local
- Sistema endocrino
Referencias
- ↑ Pandit, Nikita K. (2007). Introducción a las ciencias farmacéuticas . Lippincott Williams & Wilkins. pág . 238. ISBN 978-0-7817-4478-2.
- 1 2 Bafico, Anna; Liu, Guizhong; Goldin, Luba; Harris, Violaine; Aaronson, Stuart A. (2004). "Un mecanismo autocrino para la activación constitutiva de la vía Wnt en células cancerosas humanas" . Cancer Cell . 6 (5): 497– 506. doi : 10.1016/j.ccr.2004.09.032 . PMID 15542433 .
- 1 2 3 Schlange, Thomas; Matsuda, Yutaka; Lienhard, Susanne; Huber, Alexandre; Hynes, Nancy E (2007). "La señalización WNT autocrina contribuye a la proliferación de células de cáncer de mama a través de la vía WNT canónica y la transactivación del EGFR" . Breast Cancer Research . 9 (5): R63. doi : 10.1186/bcr1769 . PMC 2242658. PMID 17897439 .
- ↑ Grivennikov, Sergei; Karin, Michael (2008). "Señalización autocrina de IL-6: ¿un evento clave en la tumorigénesis?" . Cancer Cell . 13 (1): 7– 9. doi : 10.1016/j.ccr.2007.12.020 . PMID 18167335 .
- ↑ Gao, Sizhi Paul; Mark, Kevin G.; Leslie, Kenneth; Pao, William; Motoi, Noriko; Gerald, William L.; Travis, William D.; Bornmann, William; Veach, Darren; Clarkson, Bayard; Bromberg, Jacqueline F. (2007). "Las mutaciones en el dominio de la quinasa del EGFR median la activación de STAT3 a través de la producción de IL-6 en adenocarcinomas pulmonares humanos" . Journal of Clinical Investigation . 117 (12): 3846– 56. doi : 10.1172/JCI31871 . PMC 2096430. PMID 18060032 .
- 1 2 Hartman, Zachary C.; Yang, Xiao-Yi; Glass, Oliver; Lei, Gangjun; Osada, Takuya; Dave, Sandeep S.; Morse, Michael A.; Clay, Timothy M.; Lyerly, Herbert Kim (2011). "La sobreexpresión de HER2 desencadena un bucle de señalización autocrina proinflamatoria de IL-6 que es crítico para la tumorigénesis" . Cancer Research . 71 (13): 4380– 91. doi : 10.1158/0008-5472.CAN-11-0308 . PMC 3129398. PMID 21518778 .
- ^ Sansone, Pasquale; Storci, Gianluca; Tavolari, Simona; Guarnieri, Tiziana; Giovannini, Catia; Taffurelli, Mario; Ceccarelli, Claudio; Santini, Donatella; Paterini, Paola; Marcu, Kenneth B.; Chieco, Pasquale; Bonafè, Massimiliano (2007). "La IL-6 desencadena características malignas en mamósferas a partir del carcinoma ductal de mama humano y la glándula mamaria normal" . Revista de investigación clínica . 117 (12): 3988–4002.doi : 10.1172 / JCI32533 . PMC 2096439 . PMID 18060036 .
- ↑ Buffière A, Uzan B, Aucagne R, Hermetet F, Mas M, Nassurdine S, Aznague A, Carmignac V, Tournier B, Bouchot O, Ballerini P, Barata JT, Bastie JN, Delva L, Pflumio F, Quéré R (noviembre de 2019). "La leucemia linfoblástica aguda de células T muestra una producción autocrina de interleucina-7". Oncogén . 38 (1): 7357– 7365. doi : 10.1038/s41388-019-0921-4 . PMID 31417180 . S2CID 199668368 .
- 1 2 3 Weigand, Melanie; Hantel, Pia; Kreienberg, Rolf; Waltenberger, Johannes (2005). "Señalización autocrina del factor de crecimiento endotelial vascular en el cáncer de mama. Evidencia de líneas celulares y cultivos primarios de cáncer de mama in vitro". Angiogenesis . 8 (3): 197– 204. doi : 10.1007/s10456-005-9010-0 . PMID 16328160 . S2CID 25711624 .
- ↑ Mercurio, Arthur M; Bachelder, Robin E; Bates, Richard C; Chung, Jun (2004). "Señalización autocrina en el carcinoma: VEGF y la integrina α6β4". Seminars in Cancer Biology . 14 (2): 115– 22. doi : 10.1016/j.semcancer.2003.09.016 . PMID 15018895 .
- 1 2 Jechlinger, M.; Sommer, A; Moriggl, R; Seither, P; Kraut, N; Capodiecci, P; Donovan, M; Cordon-Cardo, C ; Beug, H; Grünert, S (2006). "La señalización autocrina de PDGFR promueve la metástasis del cáncer de mama" . Journal of Clinical Investigation . 116 (6): 1561– 70. doi : 10.1172/JCI24652 . PMC 1469776. PMID 16741576 .
- ↑ Wilson, Timothy R.; Lee, Diana Y.; Berry, Leanne; Shames, David S.; Settleman, Jeff (2011). "La señalización autocrina mediada por neuregulina-1 subyace a la sensibilidad a los inhibidores de la quinasa HER2 en un subconjunto de cánceres humanos" . Cancer Cell . 20 (2): 158–72 . doi : 10.1016/j.ccr.2011.07.011 . PMID 21840482 .
- ↑ Petersen, Sean L.; Wang, Lai; Yalcin-Chin, Asligul; Li, Lin; Peyton, Michael; Minna, John; Harran, Patrick; Wang, Xiaodong (2007). "La señalización autocrina de TNFα hace que las células cancerosas humanas sean susceptibles a la apoptosis inducida por miméticos de Smac" . Cancer Cell . 12 (5): 445– 56. doi : 10.1016/j.ccr.2007.08.029 . PMC 3431210. PMID 17996648 .
- ↑ Marek, Lindsay; Ware, Kathryn E.; Fritzsche, Alexa; Hercule, Paula; Helton, Wallace R.; Smith, Jennifer E.; McDermott, Lee A.; Coldren, Christopher D.; Nemenoff, Raphael A.; Merrick, DT; Helfrich, BA; Bunn Jr, PA; Heasley, LE (2008). "Factor de crecimiento de fibroblastos (FGF) y señalización autocrina mediada por el receptor de FGF en células de cáncer de pulmón de células no pequeñas" . Farmacología molecular . 75 (1): 196– 207. doi : 10.1124/mol.108.049544 . PMC 2669785. PMID 18849352 .
- ↑ Yi, Eun Hee; Lee, Chang Seok; Lee, Jin-Ku; Lee, Young Ju; Shin, Min Kyung; Cho, Chung-Hyun; Kang, Keon Wook; Lee, Jung Weon; Han, Wonshik; Noh, D.-Y.; Kim, Y.-N.; Cho, I.-H.; Ye, S.-k. (2012). "La señalización autocrina STAT3-RANTES es esencial para la resistencia al tamoxifeno en células de cáncer de mama humano" . Molecular Cancer Research . 11 (1): 31– 42. doi : 10.1158/1541-7786.MCR-12-0217 . PMID 23074171 .
Enlaces externos
- Señalización autocrina en los Encabezamientos de Materias Médicas (MeSH) de la Biblioteca Nacional de Medicina de EE. UU.
- "Modos de señalización autocrina versus yuxtacrina" - ilustración en sysbio.org
- transducción de señales