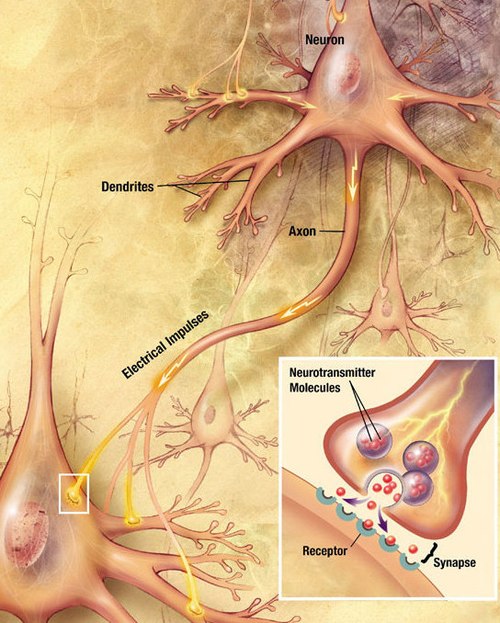
Una autapsis es una sinapsis química o eléctrica de una neurona sobre sí misma. [1] [2] También puede describirse como una sinapsis formada por el axón de una neurona sobre sus propias dendritas , in vivo o in vitro .
Historia
El término "autapsis" fue acuñado por primera vez en 1972 por Van der Loos y Glaser, quienes las observaron en preparaciones de Golgi de la corteza occipital de conejo mientras realizaban originalmente un análisis cuantitativo de los circuitos del neocórtex . [3] También en la década de 1970, se han descrito autapsis en la corteza cerebral de perros y ratas , [4] [5] [6] el neoestriado de monos , [7] y la médula espinal de gatos . [8]
En 2000, se modelaron por primera vez como soporte de la persistencia en redes neuronales recurrentes . [1] En 2004, se modelaron como demostración de comportamiento oscilatorio , que estaba ausente en la misma neurona modelo sin autapsis. [9] Más específicamente, la neurona oscilaba entre altas tasas de disparo y supresión de disparo, lo que refleja el comportamiento de ráfaga de picos que se encuentra típicamente en las neuronas cerebrales. En 2009, las autapsis se asociaron, por primera vez, con la activación sostenida. Esto propuso una posible función para las autapsis excitatorias dentro de un circuito neuronal. [10] En 2014, se demostró que las autapsis eléctricas generan ondas espirales y objetivo estables en una red modelo neuronal . [11] Esto indicó que desempeñaban un papel importante en la estimulación y regulación del comportamiento colectivo de las neuronas en la red. En 2016, se ofreció un modelo de resonancia. [12]
Las autapses se han utilizado para simular las condiciones de la "misma célula" para ayudar a los investigadores a realizar comparaciones cuantitativas, como estudiar cómo los antagonistas del receptor N -metil-D-aspartato (NMDAR) afectan a los NMDAR sinápticos frente a los extrasinápticos. [13]
Formación
Recientemente, se ha propuesto que las autapsis podrían posiblemente formarse como resultado del bloqueo de la transmisión de señales neuronales, como en casos de lesión axonal inducida por envenenamiento o impedimento de los canales iónicos. [14] Las dendritas del soma además de un axón auxiliar pueden desarrollarse para formar una autapsis para ayudar a remediar la transmisión de señales de la neurona.
Estructura y función
Las autapsis pueden ser liberadoras de glutamato (excitadoras) o liberadoras de GABA (inhibidoras), al igual que sus contrapartes sinápticas tradicionales. [15] De manera similar, las autapsis pueden ser eléctricas o químicas por naturaleza. [2]
En términos generales, la retroalimentación negativa en las autapsis tiende a inhibir las neuronas excitables, mientras que la retroalimentación positiva puede estimular las neuronas quiescentes. [16]
Aunque la estimulación de las autapsis inhibitorias no indujo potenciales postsinápticos inhibitorios hiperpolarizantes en las interneuronas de la capa V de cortes neocorticales, se ha demostrado que afectan la excitabilidad. [17] Al utilizar un antagonista de GABA para bloquear las autapsis, la probabilidad de un segundo paso de despolarización inmediatamente posterior aumentó después de un primer paso de despolarización. Esto sugiere que las autapsis actúan suprimiendo el segundo de dos pasos de despolarización estrechamente sincronizados y, por lo tanto, pueden proporcionar inhibición por retroalimentación a estas células. Este mecanismo también puede explicar potencialmente la inhibición por derivación .
En cultivos celulares, se ha demostrado que las autapsis contribuyen a la activación prolongada de las neuronas B31/B32, que contribuyen significativamente al comportamiento de respuesta a los alimentos en Aplysia . [10] Esto sugiere que las autapsis pueden desempeñar un papel en la mediación de la retroalimentación positiva. La autapsis B31/B32 no pudo desempeñar un papel en la iniciación de la actividad de la neurona, aunque se cree que ayudó a mantener el estado despolarizado de la neurona. El grado en que las autapsis mantienen la despolarización sigue sin estar claro, en particular porque otros componentes del circuito neuronal (es decir, las neuronas B63) también son capaces de proporcionar una fuerte entrada sináptica durante la despolarización. Además, se ha sugerido que las autapsis proporcionan a las neuronas B31/B32 la capacidad de repolarizarse rápidamente . Bekkers (2009) ha propuesto que bloquear específicamente la contribución de las autapsis y luego evaluar las diferencias con o sin autapsis bloqueadas podría iluminar mejor la función de las autapsis. [18]
Las neuronas del modelo Hindmarsh-Rose (HR) han demostrado patrones caóticos, de picos regulares , quiescentes y periódicos de disparos en ráfagas sin autapsis. [19] Tras la introducción de una autapsis eléctrica, el estado periódico cambia al estado caótico y muestra un comportamiento alterno que aumenta en frecuencia con una mayor intensidad autáptica y retraso temporal. Por otro lado, las autapsis químicas excitatorias mejoraron el estado caótico general. El estado caótico se redujo y suprimió en las neuronas con autapsis químicas inhibidoras. En las neuronas del modelo HR sin autapsis, el patrón de disparo cambió de quiescente a periódico y luego a caótico a medida que aumentaba la corriente CC . En general, las neuronas del modelo HR con autapsis tienen la capacidad de cambiar a cualquier patrón de disparo, independientemente del patrón de disparo anterior.
Ubicación
Se ha descubierto que las neuronas de varias regiones del cerebro, como el neocórtex, la sustancia negra y el hipocampo, contienen autapsis. [3] [20] [21] [22]
Se ha observado que las autapsis son relativamente más abundantes en las células GABAérgicas dirigidas a las dendritas y a las cestas de la corteza visual del gato en comparación con las células estrelladas espinosas , las células de doble ramo y las células piramidales , lo que sugiere que el grado de autoinervación de las neuronas es específico de cada célula. [23] Además, las autapsis de las células dirigidas a las dendritas estaban, en promedio, más alejadas del soma en comparación con las autapsis de las células de la cesta.
El 80% de las neuronas piramidales de la capa V en los neocórtex de ratas en desarrollo contenían conexiones autápticas, que se ubicaban más en las dendritas basales y las dendritas oblicuas apicales que en las dendritas apicales principales . [24] Las posiciones dendríticas de las conexiones sinápticas del mismo tipo de célula eran similares a las de las autapsis, lo que sugiere que las redes autápticas y sinápticas comparten un mecanismo común de formación.
Implicaciones de la enfermedad
En la década de 1990, se sugirió que las descargas epileptiformes interictales de tipo de cambio despolarizante paroxístico dependen principalmente de la actividad autáptica de neuronas hipocampales excitatorias solitarias de rata cultivadas en microcultivo. [25]
Más recientemente, en tejidos neocorticales humanos de pacientes con epilepsia intratable , se ha demostrado que las autapsis de salida GABAérgica de las neuronas de disparo rápido (FS) tienen una liberación asincrónica (AR) más fuerte en comparación con el tejido no epiléptico y otros tipos de sinapsis que involucran neuronas FS. [26] El estudio encontró resultados similares utilizando también un modelo de rata. Se sugirió que un aumento en la concentración residual de Ca2+ además de la amplitud del potencial de acción en las neuronas FS causa este aumento en la AR del tejido epiléptico. Los fármacos antiepilépticos podrían potencialmente dirigirse a esta AR de GABA que parece ocurrir desenfrenadamente en las autapsis de las neuronas FS.
Efectos de las drogas
Se ha demostrado que el uso de un medio acondicionado con glía para tratar microcultivos de ganglios de retina de rata purificados y libres de glía aumenta significativamente la cantidad de autapsis por neurona en comparación con un control. [27] Esto sugiere que los factores solubles derivados de la glía y sensibles a la proteinasa K inducen la formación de autapsis en las células ganglionares de la retina de rata.
Referencias
- ^ ab Seung, H. Sebastian; Lee, Daniel D.; Reis, Ben Y.; Tank, David W. (1 de septiembre de 2000). "La autapsis: una ilustración simple del almacenamiento de memoria analógica a corto plazo mediante retroalimentación sináptica ajustada". Revista de neurociencia computacional . 9 (2): 171– 185. doi :10.1023/A:1008971908649. ISSN 0929-5313. PMID 11030520. S2CID 547421.
- ^ ab Yilmaz, Ergin; Ozer, Mahmut; Baysal, Veli; Perc, Matjaž (2 de agosto de 2016). "Resonancia de coherencia múltiple inducida por autapsis en neuronas individuales y redes neuronales". Scientific Reports . 6 (1): 30914. Bibcode :2016NatSR...630914Y. doi :10.1038/srep30914. ISSN 2045-2322. PMC 4969620 . PMID 27480120.
- ^ ab Van der Loos, H.; Glaser, EM (24 de diciembre de 1972). "Autapsis en el neocórtex cerebral: sinapsis entre el axón de una célula piramidal y sus propias dendritas". Brain Research . 48 : 355– 360. doi :10.1016/0006-8993(72)90189-8. ISSN 0006-8993. PMID 4645210.
- ^ Shkol'nik-Yarros, Ekaterina G. (1971). Neuronas y conexiones interneuronales del sistema visual central | SpringerLink . doi :10.1007/978-1-4684-0715-0. ISBN 978-1-4684-0717-4. Número de identificación del sujeto 37317913.
- ^ Preston, RJ; Bishop, GA; Kitai, ST (1980-02-10). "Proyección de neuronas espinosas medianas del cuerpo estriado de la rata: un estudio intracelular de peroxidasa de rábano picante". Brain Research . 183 (2): 253– 263. doi :10.1016/0006-8993(80)90462-X. ISSN 0006-8993. PMID 7353139. S2CID 1827091.
- ^ Peters, A.; Proskauer, CC (abril de 1980). "Relaciones sinápticas entre una célula estrellada multipolar y una neurona piramidal en la corteza visual de la rata. Un estudio combinado de Golgi y microscopio electrónico". Journal of Neurocytology . 9 (2): 163– 183. doi :10.1007/bf01205156. ISSN 0300-4864. PMID 6160209. S2CID 34203892.
- ^ DiFiglia, M.; Pasik, P.; Pasik, T. (1976-09-17). "Un estudio de Golgi de los tipos neuronales en el neoestriado de los monos". Brain Research . 114 (2): 245– 256. doi :10.1016/0006-8993(76)90669-7. ISSN 0006-8993. PMID 822916. S2CID 40311354.
- ^ Scheibel, ME; Scheibel, AB (1971). "Inhibición y la célula de Renshaw: una crítica estructural; págs. 73-93". Cerebro, comportamiento y evolución . 4 (1): 73-93 . doi :10.1159/000125425. ISSN 0006-8977.
- ^ Herrmann, Christoph S. (agosto de 2004). "Autapse convierte una neurona en un oscilador". Revista internacional de bifurcación y caos . 4 (2): 623– 633. Bibcode :2004IJBC...14..623H. doi :10.1142/S0218127404009338.
- ^ ab Saada, R.; Miller, N.; Hurwitz, I.; Susswein, AJ (2009). "La excitación muscarínica autáptica subyace a un potencial de meseta y a una actividad persistente en una neurona de función conductual conocida". Current Biology . 19 (6): 479– 484. doi : 10.1016/j.cub.2009.01.060 . PMID 19269179. S2CID 15990017.
- ^ Qin, H.; Ma, J.; Wang, C.; Chu, R. (2014). "Onda objetivo inducida por autapsis, onda espiral en red regular de neuronas". Science China Physics, Mechanics & Astronomy . 57 (10): 1918– 1926. Bibcode :2014SCPMA..57.1918Q. doi :10.1007/s11433-014-5466-5. S2CID 120661751.
- ^ Yilmaz, E.; Ozer, M.; Baysal, V.; Perc, M. (2 de agosto de 2016). "Resonancia de coherencia múltiple inducida por autapsis en neuronas individuales y redes neuronales". Scientific Reports . 9 : 30914. Bibcode :2016NatSR...630914Y. doi :10.1038/srep30914. PMC 4969620 . PMID 27480120.
- ^ Xia, Peng; Chen, Huei-sheng Vincent; Zhang, Dongxian; Lipton, Stuart A. (18 de agosto de 2010). "La memantina bloquea preferentemente las corrientes extrasinápticas sobre las corrientes sinápticas del receptor NMDA en las autapsis del hipocampo". Journal of Neuroscience . 30 (33): 11246– 11250. doi :10.1523/JNEUROSCI.2488-10.2010. ISSN 0270-6474. PMC 2932667 . PMID 20720132.
- ^ Wang, Chunni; Guo, Shengli; Xu, Ying; Mamá, junio; Tang, junio; Alzahrani, Faris; Hobiny, Aatef (2017). "Formación de autapsis conectada a la neurona y su función biológica". Complejidad . 2017 : 1– 9. doi : 10.1155/2017/5436737 . ISSN 1076-2787.
- ^ Ikeda, Kaori; Bekkers, John M. (9 de mayo de 2006). "Autapses". Current Biology . 16 (9): R308. Bibcode :2006CBio...16.R308I. doi : 10.1016/j.cub.2006.03.085 . ISSN 0960-9822. PMID 16682332.
- ^ Qin, Huixin; Wu, Ying; Wang, Chunni; Ma, Jun (2015). "Emisión de ondas desde defectos en la red con autapses". Comunicaciones en ciencia no lineal y simulación numérica . 23 ( 1–3 ): 164–174 . Bibcode :2015CNSNS..23..164Q. doi :10.1016/j.cnsns.2014.11.008.
- ^ Bacci, Alberto; Huguenard, John R.; Prince, David A. (1 de febrero de 2003). "Neurotransmisión autáptica funcional en interneuronas de activación rápida: una nueva forma de inhibición por retroalimentación en el neocórtex". The Journal of Neuroscience . 23 (3): 859– 866. doi : 10.1523/JNEUROSCI.23-03-00859.2003 . ISSN 1529-2401. PMC 6741939 . PMID 12574414.
- ^ Bekkers, John M. (2009). "Transmisión sináptica: ¿Las autapsis excitatorias encuentran una función?". Current Biology . 19 (7): R296 – R298 . Bibcode :2009CBio...19.R296B. doi : 10.1016/j.cub.2009.02.010 . PMID 19368875. S2CID 15821336.
- ^ Wang, Hengtong; Ma, Jun; Chen, Yueling; Chen, Yong (2014). "Efecto de una autapsis en la transición del patrón de disparo en una neurona en ráfaga". Communications in Nonlinear Science and Numerical Simulation . 19 (9): 3242– 3254. Bibcode :2014CNSNS..19.3242W. doi :10.1016/j.cnsns.2014.02.018.
- ^ Park, Melburn R.; Lighthall, James W.; Kitai, Stephen T. (1980). "Inhibición recurrente en el neoestriado de la rata". Brain Research . 194 (2): 359– 369. doi :10.1016/0006-8993(80)91217-2. PMID 7388619. S2CID 29451737.
- ^ Karabelas, Athanasios B.; Purrura, Dominick P. (1980). "Evidencia de autapsis en la sustancia negra". Brain Research . 200 (2): 467– 473. doi :10.1016/0006-8993(80)90935-x. PMID 6158366. S2CID 35517474.
- ^ Cobb, SR; Halasy, K; Vida, I; Nyı́ri, G; Tamás, G; Buhl, EH; Somogyi, P (1997). "Efectos sinápticos de las interneuronas identificadas que inervan tanto a las interneuronas como a las células piramidales en el hipocampo de la rata". Neurociencia . 79 (3): 629– 648. doi :10.1016/s0306-4522(97)00055-9. PMID 9219929. S2CID 15479304.
- ^ Tamás, G.; Buhl, EH; Somogyi, P. (15 de agosto de 1997). "Autoinervación autáptica masiva de neuronas GABAérgicas en la corteza visual del gato". The Journal of Neuroscience . 17 (16): 6352– 6364. doi : 10.1523/JNEUROSCI.17-16-06352.1997 . ISSN 0270-6474. PMC 6568358 . PMID 9236244.
- ^ Lübke, J.; Markram, H.; Frotscher, M.; Sakmann, B. (15 de mayo de 1996). "Frecuencia y distribución dendrítica de las autapsis establecidas por neuronas piramidales de capa 5 en el neocórtex de rata en desarrollo: comparación con la inervación sináptica de neuronas adyacentes de la misma clase". The Journal of Neuroscience . 16 (10): 3209– 3218. doi : 10.1523/JNEUROSCI.16-10-03209.1996 . ISSN 0270-6474. PMC 6579140 . PMID 8627359.
- ^ Segal, MM (octubre de 1994). "Las explosiones endógenas subyacen a la actividad similar a una convulsión en neuronas hipocampales excitatorias solitarias en microcultivos". Journal of Neurophysiology . 72 (4): 1874– 1884. doi :10.1152/jn.1994.72.4.1874. ISSN 0022-3077. PMID 7823106.
- ^ Jiang, hombre; Zhu, Jie; Liu, Yaping; Yang, Mingpo; Tian, Cuiping; Jiang, Shan; Wang, Yonghong; Guo, Hui; Wang, Kaiyan (8 de mayo de 2012). "Mejora de la liberación asincrónica de interneuronas de pico rápido en la neocorteza epiléptica humana y de rata". Más biología . 10 (5): e1001324. doi : 10.1371/journal.pbio.1001324 . ISSN 1545-7885. PMC 3348166 . PMID 22589699.
- ^ Nägler, Karl; Mauch, Daniela H; Pfrieger, Frank W (15 de junio de 2001). "Las señales derivadas de la glía inducen la formación de sinapsis en neuronas del sistema nervioso central de la rata". The Journal of Physiology . 533 (Pt 3): 665– 679. doi :10.1111/j.1469-7793.2001.00665.x. ISSN 0022-3751. PMC 2278670 . PMID 11410625.