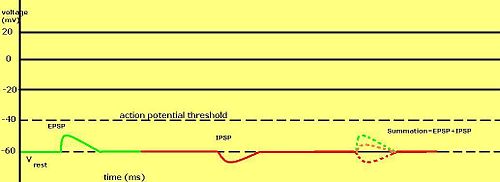
Un potencial postsináptico inhibitorio ( PPSI ) es un tipo de potencial sináptico que hace que una neurona postsináptica sea menos propensa a generar un potencial de acción . [ 1 ] Lo opuesto a un potencial postsináptico inhibitorio es un potencial postsináptico excitatorio (PPSE), que es un potencial sináptico que hace que una neurona postsináptica sea más propensa a generar un potencial de acción. Los PPSI pueden ocurrir en todas las sinapsis químicas, que utilizan la secreción de neurotransmisores para crear señalización de célula a célula. Los PPSE y los PPSI compiten entre sí en numerosas sinapsis de una neurona. Esto determina si un potencial de acción que ocurre en el terminal presináptico produce un potencial de acción en la membrana postsináptica. Algunos neurotransmisores comunes involucrados en los PPSI son GABA y glicina .
Las neuronas presinápticas inhibitorias liberan neurotransmisores que se unen a los receptores postsinápticos ; esto induce un cambio en la permeabilidad de la membrana neuronal postsináptica a iones específicos. Se genera una corriente eléctrica que modifica el potencial de membrana postsináptica, creando un potencial postsináptico más negativo ; es decir, el potencial de membrana postsináptica se vuelve más negativo que el potencial de membrana en reposo, y esto se denomina hiperpolarización . Para generar un potencial de acción, la membrana postsináptica debe despolarizarse ; el potencial de membrana debe alcanzar un umbral de voltaje más positivo que el potencial de membrana en reposo. Por lo tanto, la hiperpolarización de la membrana postsináptica reduce la probabilidad de que se produzca una despolarización suficiente para generar un potencial de acción en la neurona postsináptica.
La despolarización también puede ocurrir debido a un IPSP si el potencial inverso está entre el umbral de reposo y el umbral del potencial de acción . Otra forma de ver los potenciales postsinápticos inhibitorios es que también son un cambio en la conductancia de cloruro en la célula neuronal porque disminuye la fuerza impulsora. [ 2 ] Esto se debe a que, si el neurotransmisor liberado en la hendidura sináptica causa un aumento en la permeabilidad de la membrana postsináptica a los iones cloruro al unirse a los canales iónicos de cloruro regulados por ligando y hacer que se abran, entonces los iones cloruro, que están en mayor concentración en la hendidura sináptica, se difunden en la neurona postsináptica. Como estos son iones con carga negativa, se produce una hiperpolarización, lo que hace menos probable que se genere un potencial de acción en la neurona postsináptica. Se pueden usar microelectrodos para medir potenciales postsinápticos tanto en sinapsis excitatorias como inhibitorias.
En general, un potencial postsináptico depende del tipo y la combinación del canal receptor, el potencial inverso del potencial postsináptico, el voltaje umbral del potencial de acción , la permeabilidad iónica del canal iónico, así como las concentraciones de iones dentro y fuera de la célula; esto determina si es excitatorio o inhibitorio. Los potenciales postsinápticos inhibitorios (PPSI) siempre tienden a mantener el potencial de membrana más negativo que el umbral del potencial de acción y pueden considerarse como una "hiperpolarización transitoria". [ 3 ]
Los IPSP fueron investigados por primera vez en motoneuronas por David PC Lloyd, John Eccles y Rodolfo Llinás en las décadas de 1950 y 1960. [ 4 ] [ 5 ]

Componentes
Tipos
Este sistema [ 1 ] permite que los IPSP se sumen temporalmente con los EPSP subumbrales o supraumbrales para reducir la amplitud del potencial postsináptico resultante. Los EPSP (positivos) y los IPSP (negativos) equivalentes se anulan entre sí al sumarse. El equilibrio entre los EPSP y los IPSP es crucial para la integración de la información eléctrica producida por las sinapsis inhibitorias y excitatorias.

Factores
El tamaño de la neurona también puede afectar el potencial postsináptico inhibitorio. En las neuronas más pequeñas se produce una simple sumación temporal de los potenciales postsinápticos, mientras que en las neuronas más grandes, un mayor número de sinapsis y receptores ionotrópicos, así como una mayor distancia entre la sinapsis y el soma, permiten la prolongación de las interacciones entre neuronas.
Neurotransmisores inhibidores
El GABA es un neurotransmisor muy común utilizado en los IPSP en el cerebro y la retina de los mamíferos adultos. [ 1 ] [ 6 ] Las moléculas de glicina y sus receptores funcionan de manera muy similar en la médula espinal, el cerebro y la retina.
Receptores inhibidores
Existen dos tipos de receptores inhibidores:
receptores ionotrópicos
Los receptores ionotrópicos (también conocidos como canales iónicos regulados por ligando) desempeñan un papel importante en los potenciales postsinápticos inhibitorios. [ 1 ] Un neurotransmisor se une al sitio extracelular y abre el canal iónico, que consta de un dominio transmembrana que permite el paso de iones a través de la membrana dentro de la célula postsináptica. Este tipo de receptor produce acciones postsinápticas muy rápidas, apenas unos milisegundos después de que la terminal presináptica reciba un potencial de acción. Estos canales influyen en la amplitud y la duración de los potenciales postsinápticos en su conjunto.
Los receptores ionotrópicos de GABA ( receptores GABA A ) son pentámeros compuestos generalmente por tres subunidades diferentes (α, β, γ), aunque existen otras subunidades (δ, ε, θ, π, ρ) y conformaciones. Los canales abiertos son selectivamente permeables a los iones cloruro o potasio (según el tipo de receptor) y permiten que estos iones atraviesen la membrana. Si el potencial electroquímico del ion es más negativo que el umbral del potencial de acción, el cambio de conductancia resultante de la unión del GABA a sus receptores mantiene el potencial postsináptico más negativo que el umbral y disminuye la probabilidad de que la neurona postsináptica complete un potencial de acción.
Los receptores ionotrópicos de GABA se utilizan para la unión de diversos fármacos, como barbitúricos ( fenobarbital , pentobarbital ), esteroides y picrotoxina . Las benzodiazepinas (Valium) se unen a las subunidades α y γ de los receptores GABA para mejorar la señalización gabaérgica. El alcohol también modula los receptores ionotrópicos de GABA.
Receptores metabotrópicos
Los receptores metabotrópicos suelen ser receptores acoplados a proteínas G , como los receptores GABA B. Estos no utilizan canales iónicos en su estructura; en cambio, constan de un dominio extracelular que se une a un neurotransmisor y un dominio intracelular que se une a la proteína G. [ 1 ] Esto inicia la activación de la proteína G, que luego se libera del receptor e interactúa con canales iónicos y otras proteínas para abrir o cerrar canales iónicos a través de mensajeros intracelulares. Producen respuestas postsinápticas lentas (de milisegundos a minutos) y pueden activarse junto con receptores ionotrópicos para crear potenciales postsinápticos rápidos y lentos en una sinapsis particular. Los receptores metabotrópicos GABA, heterodímeros de subunidades R1 y R2, utilizan canales de potasio en lugar de cloruro. También pueden bloquear los canales de iones de calcio para hiperpolarizar las células postsinápticas.
Significado
Los potenciales postsinápticos inhibitorios tienen numerosas aplicaciones en el mundo real. Los fármacos que afectan la acción de los neurotransmisores pueden tratar trastornos neurológicos y psicológicos mediante diferentes combinaciones de tipos de receptores, proteínas G y canales iónicos en las neuronas postsinápticas.
Por ejemplo, se están realizando estudios que investigan la desensibilización y el tráfico de receptores opioides en el locus coeruleus del cerebro. Cuando se aplica una alta concentración de agonista durante un tiempo prolongado (quince minutos o más), la hiperpolarización alcanza su punto máximo y luego disminuye. Esto es significativo porque es un preludio a la tolerancia; cuantos más opioides se necesiten para el dolor, mayor será la tolerancia del paciente. Estos estudios son importantes porque nos ayudan a comprender mejor cómo afrontamos el dolor y nuestras respuestas a diversas sustancias que ayudan a tratarlo. Al estudiar nuestra tolerancia al dolor, podemos desarrollar medicamentos más eficaces para su tratamiento. [ 7 ]
Además, se están realizando investigaciones en el campo de las neuronas dopaminérgicas en el área tegmental ventral, que se relaciona con la recompensa, y la sustancia negra, que está involucrada en el movimiento y la motivación. Las respuestas metabotrópicas ocurren en las neuronas dopaminérgicas a través de la regulación de la excitabilidad celular. Los opioides inhiben la liberación de GABA; esto disminuye la cantidad de inhibición y les permite disparar espontáneamente. La morfina y los opioides se relacionan con potenciales postsinápticos inhibitorios porque inducen desinhibición en las neuronas dopaminérgicas. [ 7 ]
Los IPSP también pueden usarse para estudiar las características de entrada-salida de una sinapsis inhibitoria del prosencéfalo utilizada para estudiar más a fondo el comportamiento aprendido, por ejemplo, en un estudio de aprendizaje de canto en aves en la Universidad de Washington. [ 8 ] Se indujeron trenes de Poisson de IPSP unitarios a alta frecuencia para reproducir el disparo postsináptico en la porción medial del núcleo talámico dorsalateral sin ninguna entrada excitatoria adicional. Esto muestra un exceso de activación GABAérgica talámica. Esto es importante porque la sincronización del disparo es necesaria para la correcta localización del sonido en las vías auditivas ascendentes. Las aves cantoras usan terminales sinápticas caliciformes GABAérgicas y una sinapsis similar a un cáliz de modo que cada célula en el núcleo talámico dorsalateral recibe como máximo dos terminales axónicos de los ganglios basales para crear grandes corrientes postsinápticas.
Los potenciales postsinápticos inhibitorios también se utilizan para estudiar los ganglios basales de los anfibios para ver cómo se modula la función motora a través de sus salidas inhibitorias del estriado al tectum y tegmento. [ 9 ] Los comportamientos guiados visualmente pueden estar regulados a través de la vía estriato-tegmental inhibitoria encontrada en anfibios en un estudio realizado en el Baylor College of Medicine y la Academia China de Ciencias. Los ganglios basales en los anfibios son muy importantes para recibir entradas visuales, auditivas, olfativas y mecanosensoriales; la vía estriato-protecto-tectal desinhibitoria es importante en los comportamientos de captura de presas de los anfibios. Cuando el estriado ipsilateral de un sapo adulto fue estimulado eléctricamente, se indujeron potenciales postsinápticos inhibitorios en neuronas tegmentales binoculares, lo que afecta el sistema visual del sapo.
Estudios
Los potenciales postsinápticos inhibitorios pueden ser inhibidos a través de un proceso de señalización llamado " supresión de la inhibición inducida por despolarización (DSI)" en las células piramidales CA1 y las células de Purkinje cerebelosas. [ 10 ] [ 11 ] En un entorno de laboratorio, se han utilizado despolarizaciones escalonadas del soma para crear DSI, pero también puede lograrse a través de la despolarización sináptica inducida de las dendritas. Las DSI pueden ser bloqueadas por antagonistas de los canales iónicos de calcio del receptor ionotrópico en los somas y las dendritas apicales proximales de las células piramidales CA1. Los potenciales postsinápticos inhibitorios dendríticos pueden ser severamente reducidos por las DSI a través de la despolarización directa.
En este sentido, los potenciales postsinápticos inhibitorios son útiles en la señalización del bulbo olfatorio a la corteza olfativa . [ 12 ] Los EPSP se amplifican por la conductancia persistente de iones de sodio en las células en penacho externas . La conductancia de iones de calcio activada por bajo voltaje mejora aún más los EPSP. La conductancia catiónica no selectiva activada por hiperpolarización disminuye la sumación y la duración de los EPSP y también cambia las entradas inhibitorias a la excitación postsináptica. Los IPSP entran en escena cuando las membranas de las células en penacho se despolarizan y los IPSP causan entonces inhibición. En el umbral de reposo, los IPSP inducen potenciales de acción. El GABA es responsable de gran parte del trabajo de los IPSP en las células en penacho externas.
Otro estudio interesante de potenciales postsinápticos inhibitorios examina las oscilaciones del ritmo theta neuronal que pueden usarse para representar fenómenos electrofisiológicos y diversos comportamientos. [ 13 ] [ 14 ] Los ritmos theta se encuentran en el hipocampo y la inhibición sináptica GABAérgica ayuda a modularlos. Son dependientes de los IPSP y se inician en CA3 por receptores muscarínicos de acetilcolina y dentro de C1 por la activación de receptores metabotrópicos de glutamato del grupo I. Cuando las interneuronas son activadas por receptores metabotrópicos de acetilcolina en la región CA1 de cortes de hipocampo de rata, se produce un patrón theta de IPSP en las células piramidales independientemente de la entrada. Esta investigación también estudia las DSI, mostrando que las DSI interrumpen el ritmo iniciado por la acetilcolina metabotrópica a través de la liberación de endocannabinoides. Un mecanismo dependiente de endocannabinoides puede interrumpir los IPSP theta a través de potenciales de acción emitidos como un patrón de ráfaga o tren breve. Además, la activación de los receptores metabotrópicos de glutamato elimina cualquier actividad IPSP theta a través de una vía independiente de iones de calcio y mediada por proteína G.
También se han estudiado los potenciales postsinápticos inhibitorios en la célula de Purkinje a través de la amplificación dendrítica. El estudio se centró en la propagación de los IPSP a lo largo de las dendritas y su dependencia de los receptores ionotrópicos mediante la medición de la amplitud y la evolución temporal del potencial postsináptico inhibitorio. Los resultados mostraron que tanto los potenciales postsinápticos inhibitorios compuestos como los unitarios se amplifican por los canales iónicos de calcio dendríticos. El ancho de un IPSP somático es independiente de la distancia entre el soma y la sinapsis, mientras que el tiempo de ascenso aumenta con esta distancia. Estos IPSP también regulan los ritmos theta en las células piramidales. Por otro lado, los potenciales postsinápticos inhibitorios son despolarizantes y a veces excitatorios en las neuronas espinales de mamíferos inmaduras debido a las altas concentraciones de cloruro intracelular a través de los canales iónicos ionotrópicos de GABA o cloruro de glicina. [ 15 ] Estas despolarizaciones activan los canales de calcio dependientes de voltaje. Posteriormente se vuelven hiperpolarizantes a medida que el mamífero madura. En concreto, en ratas, esta maduración se produce durante el periodo perinatal, cuando las proyecciones del tronco encefálico alcanzan la médula espinal lumbar. Las señales moduladoras descendentes son necesarias para el cambio evolutivo de potenciales postsinápticos inhibitorios despolarizantes a hiperpolarizantes. Esto se estudió mediante transecciones completas de la médula espinal al nacer las ratas y el registro de potenciales postsinápticos inhibitorios (PPSI) de las motoneuronas lumbares al final de la primera semana después del nacimiento.
El glutamato , un neurotransmisor excitatorio, se asocia generalmente con potenciales postsinápticos excitatorios en la transmisión sináptica. Sin embargo, un estudio realizado en el Instituto Vollum de la Universidad de Ciencias de la Salud de Oregón demuestra que el glutamato también puede utilizarse para inducir potenciales postsinápticos inhibitorios en las neuronas. [ 16 ] Este estudio explica que los receptores metabotrópicos de glutamato presentan proteínas G activadas en las neuronas dopaminérgicas que inducen la hidrólisis de fosfoinosítidos. Los productos resultantes se unen a los receptores de inositol trifosfato (IP3) a través de canales de iones de calcio. El calcio proviene de los depósitos y activa la conductancia de potasio, lo que provoca una inhibición pura en las células dopaminérgicas. Los cambios en los niveles de glutamato liberado sinápticamente crean una excitación mediante la activación de receptores ionotrópicos, seguida de la inhibición de los receptores metabotrópicos de glutamato.
Véase también
Referencias
- 1 2 3 4 5 Purves et al. Neurociencia. 4.ª ed. Sunderland (MA): Sinauer Associates, Incorporated; 2008.
- ↑ Thompson SM, Gähwiler BH (marzo de 1989). "Desinhibición dependiente de la actividad. I. La estimulación repetitiva reduce la fuerza impulsora y la conductancia del IPSP en el hipocampo in vitro". Journal of Neurophysiology . 61 (3): 501– 11. doi : 10.1152/jn.1989.61.3.501 . PMID 2709096 .
- ↑ Levy M, Koeppen B, Stanton B (2005). Principios de fisiología de Berne y Levy (4.ª ed.). Elsevier Mosby. ISBN 978-0-8089-2321-3.
- ↑ Coombs JS, Eccles JC, Fatt P (noviembre de 1955). "Las conductancias iónicas específicas y los movimientos iónicos a través de la membrana de la motoneurona que producen el potencial postsináptico inhibitorio" . The Journal of Physiology . 130 (2): 326–74 . doi : 10.1113/jphysiol.1955.sp005412 . PMC 1363415. PMID 13278905 .
- ↑ Llinas R, Terzuolo CA (marzo de 1965). "Mecanismos de acciones supraespinales sobre las actividades de la médula espinal. Mecanismos inhibitorios reticulares sobre las motoneuronas flexoras". Journal of Neurophysiology . 28 (2): 413–22 . doi : 10.1152/jn.1965.28.2.413 . PMID 14283063 .
- ↑ Chavas J, Marty A (marzo de 2003). "Coexistencia de sinapsis GABA excitatorias e inhibitorias en la red de interneuronas cerebelosas" . The Journal of Neuroscience . 23 (6): 2019–31 . doi : 10.1523/JNEUROSCI.23-06-02019.2003 . PMC 6742031. PMID 12657660 .
- 1 2 Williams, JT, Instituto Vollum de la Universidad de Ciencias de la Salud de Oregón, Entrevistado por Saira Ahmed, 11 de noviembre de 2008
- ↑ Person AL, Perkel DJ (abril de 2005). "Los IPSP unitarios impulsan la actividad talámica precisa en un circuito necesario para el aprendizaje" . Neuron . 46 (1): 129–40 . doi : 10.1016/j.neuron.2004.12.057 . PMID 15820699 .
- ↑ Wu GY, Wang SR (diciembre de 2007). "Potenciales postsinápticos y proyecciones axonales de neuronas tegmentales que responden a la estimulación eléctrica del cuerpo estriado del sapo" . Neuroscience Letters . 429 ( 2–3 ): 111–4 . doi : 10.1016/j.neulet.2007.09.071 . PMC 2696233. PMID 17996369 .
- ↑ Morishita W, Alger BE (enero de 2001). "La despolarización directa y los potenciales de acción antidrómicos suprimen transitoriamente los IPSP dendríticos en las células piramidales CA1 del hipocampo" . Journal of Neurophysiology . 85 (1): 480–4 . doi : 10.1152/jn.2001.85.1.480 . PMID 11152751. S2CID 17060042 .
- ↑ Solinas SM, Maex R, De Schutter E (marzo de 2006). "Amplificación dendrítica de potenciales postsinápticos inhibitorios en una célula de Purkinje modelo" ( PDF) . The European Journal of Neuroscience . 23 (5): 1207–18 . doi : 10.1111/j.1460-9568.2005.04564.x . PMID 16553783. S2CID 6139806. Archivado del original (PDF) el 18 de abril de 2007. Recuperado el 22 de septiembre de 2019 .
- ↑ Liu S, Shipley MT (octubre de 2008). "Las conductancias intrínsecas dan forma activamente a las respuestas postsinápticas excitatorias e inhibitorias en las células en penacho externas del bulbo olfatorio" . The Journal of Neuroscience . 28 (41): 10311– 22. doi : 10.1523/JNEUROSCI.2608-08.2008 . PMC 2570621. PMID 18842890 .
- ↑ Reich CG, Karson MA, Karnup SV, Jones LM, Alger BE (diciembre de 2005). "Regulación del ritmo theta de los potenciales postsinápticos inhibitorios por receptores muscarínicos y endocannabinoides en el hipocampo" ( PDF) . Journal of Neurophysiology . 94 (6): 4290–9 . doi : 10.1152/jn.00480.2005 . PMID 16093334. S2CID 10333266. Archivado del original (PDF) el 27 de febrero de 2019.
- ↑ Brenowitz SD, Regehr WG (2003). "Dependencia del calcio de la inhibición retrógrada por endocannabinoides en las sinapsis de las células de Purkinje" . Journal of Neuroscience . 23 (15): 6373– 6384. doi : 10.1523/JNEUROSCI.23-15-06373.2003 . PMC 6740543. PMID 12867523 .
- ↑ Jean-Xavier C, Pflieger JF, Liabeuf S, Vinay L (noviembre de 2006). "Los potenciales postsinápticos inhibitorios en las motoneuronas lumbares permanecen despolarizantes después de la transección de la médula espinal neonatal en la rata". Journal of Neurophysiology . 96 (5): 2274–81 . CiteSeerX 10.1.1.326.1283 . doi : 10.1152/jn.00328.2006 . PMID 16807348 .
- ↑ Fiorillo CD, Williams JT (julio de 1998). "El glutamato media un potencial postsináptico inhibitorio en las neuronas dopaminérgicas". Nature . 394 (6688): 78– 82. Bibcode : 1998Natur.394...78F . doi : 10.1038/27919 . PMID 9665131. S2CID 4352019 .
- Memoria
- Sinapsis neuronal
- Potenciales graduados