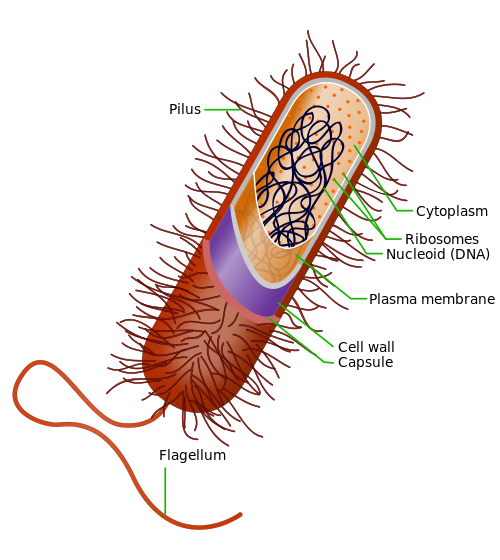
La región codificante de un gen , también conocida como secuencia codificante de ADN ( CDS ), es la porción del ADN o ARN de un gen que codifica una proteína . [ 1 ] Estudiar la longitud, composición, regulación, empalme, estructuras y funciones de las regiones codificantes en comparación con las regiones no codificantes en diferentes especies y períodos de tiempo puede proporcionar una cantidad significativa de información importante sobre la organización genética y la evolución de procariotas y eucariotas . [ 2 ] Esto puede ayudar aún más a mapear el genoma humano y desarrollar terapia génica. [ 3 ]
Definición
Aunque este término a veces se usa indistintamente con exón , no es exactamente lo mismo: el exón puede estar compuesto tanto por la región codificante como por las regiones no codificantes 3' y 5' del ARN, por lo que un exón estaría parcialmente formado por la región codificante. Las regiones no codificantes 3' y 5' del ARN, que no codifican proteínas, se denominan regiones no codificantes y no se tratan en esta página. [ 4 ]
A menudo se confunde el término regiones codificantes con el de exomas , pero existe una clara distinción entre ellos. Mientras que el exoma se refiere a todos los exones de un genoma, la región codificante se refiere a secciones del ADN (o transcrito primario ) o a una sección específica de ARNm procesado que codifica un tipo determinado de proteína.
Historia
En 1978, Walter Gilbert publicó "Por qué los genes están fragmentados", obra que comenzó a explorar la idea de que el gen es un mosaico: que cada cadena completa de ácido nucleico no está codificada de forma continua, sino que está interrumpida por regiones no codificantes "silenciosas". Esta fue la primera indicación de que era necesario distinguir entre las partes del genoma que codifican proteínas, ahora llamadas regiones codificantes, y las que no. [ 5 ]
Composición
La evidencia sugiere que existe una interdependencia general entre los patrones de composición de bases y la disponibilidad de la región codificante. [ 6 ] Se cree que la región codificante contiene un mayor contenido de GC que las regiones no codificantes. Investigaciones posteriores han descubierto que cuanto más larga es la cadena codificante, mayor es el contenido de GC. Las cadenas codificantes cortas son comparativamente pobres en GC, de forma similar al bajo contenido de GC de los codones de parada traslacionales de composición de bases como TAG, TAA y TGA. [ 7 ]
Las regiones ricas en GC también presentan una ligera alteración en la proporción de tipos de mutaciones puntuales : hay más transiciones , que son cambios de purina a purina o de pirimidina a pirimidina, en comparación con las transversiones , que son cambios de purina a pirimidina o de pirimidina a purina. Es menos probable que las transiciones modifiquen el aminoácido codificado y permanecen como mutaciones silenciosas (especialmente si ocurren en el tercer nucleótido de un codón), lo cual suele ser beneficioso para el organismo durante la traducción y la formación de proteínas. [ 8 ]
Esto indica que las regiones codificantes esenciales (ricas en genes) tienen un mayor contenido de GC y son más estables y resistentes a la mutación en comparación con las regiones accesorias y no esenciales (pobres en genes). [ 9 ] Sin embargo, aún no está claro si esto se produjo a través de una mutación neutra y aleatoria o a través de un patrón de selección . [ 10 ] También existe un debate sobre si los métodos utilizados, como las ventanas genéticas, para determinar la relación entre el contenido de GC y la región codificante son precisos e imparciales. [ 11 ]
Estructura y función

En el ADN , la región codificante está flanqueada por la secuencia promotora en el extremo 5' de la cadena molde y la secuencia de terminación en el extremo 3'. Durante la transcripción , la ARN polimerasa (ARNp) se une a la secuencia promotora y se desplaza a lo largo de la cadena molde hasta la región codificante. La ARNp añade entonces nucleótidos de ARN complementarios a la región codificante para formar el ARNm , sustituyendo la timina por uracilo . [ 12 ] Este proceso continúa hasta que la ARNp alcanza la secuencia de terminación. [ 12 ]
Después de la transcripción y maduración, el ARNm maduro formado comprende múltiples partes importantes para su eventual traducción a proteína . La región codificante en un ARNm está flanqueada por la región no traducida 5' (5'-UTR) y la región no traducida 3' (3'-UTR), [ 1 ] la caperuza 5' y la cola de poli-A . Durante la traducción , el ribosoma facilita la unión de los ARNt a la región codificante, 3 nucleótidos a la vez ( codones ). [ 13 ] Los ARNt transfieren sus aminoácidos asociados a la cadena polipeptídica en crecimiento, formando finalmente la proteína definida en la región codificante de ADN inicial.

Regulación
La región codificante puede modificarse para regular la expresión génica.
La alquilación es una forma de regulación de la región codificante. [ 15 ] El gen que se habría transcrito puede silenciarse al dirigirse a una secuencia específica. Las bases en esta secuencia se bloquearían mediante grupos alquilo , lo que crea el efecto de silenciamiento . [ 16 ]
Si bien la regulación de la expresión génica controla la abundancia de ARN o proteínas producidas en una célula, la regulación de estos mecanismos puede controlarse mediante una secuencia reguladora ubicada antes del inicio del marco de lectura abierto en una cadena de ADN. Dicha secuencia reguladora determinará entonces la ubicación y el momento en que se producirá la expresión de una región codificante de proteínas. [ 17 ]
El empalme de ARN determina en última instancia qué parte de la secuencia se traduce y expresa, y este proceso implica cortar intrones y unir exones. Sin embargo, el lugar donde corta el espliceosoma de ARN está guiado por el reconocimiento de los sitios de empalme , en particular el sitio de empalme 5', que es uno de los sustratos para el primer paso del empalme. [ 18 ] Las regiones codificantes están dentro de los exones, que se unen covalentemente para formar el ARN mensajero maduro .
Mutaciones
Las mutaciones en la región codificante pueden tener efectos muy diversos en el fenotipo del organismo. Si bien algunas mutaciones en esta región del ADN/ARN pueden resultar en cambios beneficiosos, otras pueden ser perjudiciales e incluso letales para la supervivencia del organismo. Por el contrario, los cambios en la región no codificante no siempre producen cambios detectables en el fenotipo.
Tipos de mutación

Existen diversas formas de mutaciones que pueden ocurrir en las regiones codificantes. Una forma son las mutaciones silenciosas , en las que un cambio en los nucleótidos no produce ningún cambio en el aminoácido después de la transcripción y la traducción. [ 20 ] También existen mutaciones sin sentido , donde las alteraciones de bases en la región codificante codifican un codón de parada prematuro, produciendo una proteína final más corta. Las mutaciones puntuales , o cambios de un solo par de bases en la región codificante, que codifican diferentes aminoácidos durante la traducción, se denominan mutaciones de sentido erróneo . Otros tipos de mutaciones incluyen mutaciones de cambio de marco de lectura, como inserciones o deleciones . [ 20 ]
Formación
Algunas formas de mutaciones son hereditarias ( mutaciones de la línea germinal ), es decir, se transmiten de un progenitor a su descendencia. [ 21 ] Estas regiones codificantes mutadas están presentes en todas las células del organismo. Otras formas de mutaciones se adquieren ( mutaciones somáticas ) durante la vida del organismo y pueden no ser constantes de célula a célula. [ 21 ] Estos cambios pueden ser causados por mutágenos , carcinógenos u otros agentes ambientales (por ejemplo, radiación UV ). Las mutaciones adquiridas también pueden ser resultado de errores de copia durante la replicación del ADN y no se transmiten a la descendencia. Los cambios en la región codificante también pueden ser de novo (nuevos); se cree que estos cambios ocurren poco después de la fecundación , lo que resulta en una mutación presente en el ADN de la descendencia, pero ausente tanto en los espermatozoides como en los óvulos. [ 21 ]
Prevención
Existen múltiples mecanismos de transcripción y traducción para prevenir la letalidad debida a mutaciones deletéreas en la región codificante. Dichas medidas incluyen la corrección de errores por parte de algunas ADN polimerasas durante la replicación, la reparación de errores de apareamiento después de la replicación, [ 22 ] y la « Hipótesis del Tambaleo », que describe la degeneración de la tercera base dentro de un codón de ARNm. [ 23 ]
Regiones de codificación restringidas (RCR)
Si bien es bien sabido que el genoma de un individuo puede tener diferencias extensas en comparación con el genoma de otro, investigaciones recientes han descubierto que algunas regiones codificantes están altamente restringidas, o son resistentes a la mutación, entre individuos de la misma especie. Esto es similar al concepto de restricción interespecífica en secuencias conservadas . Los investigadores denominaron a estas secuencias altamente restringidas regiones codificantes restringidas (RCR) y también descubrieron que dichas regiones pueden estar involucradas en una alta selección purificadora . En promedio, hay aproximadamente 1 mutación que altera la proteína cada 7 bases codificantes, pero algunas RCR pueden tener más de 100 bases en la secuencia sin mutaciones que alteren la proteína observadas, algunas incluso sin mutaciones sinónimas. [ 24 ] Estos patrones de restricción entre genomas pueden proporcionar pistas sobre las fuentes de enfermedades raras del desarrollo o potencialmente incluso letalidad embrionaria. Las variantes clínicamente validadas y las mutaciones de novo en las RCR se han vinculado previamente con trastornos como la encefalopatía epiléptica infantil , el retraso del desarrollo y la cardiopatía grave. [ 24 ]
Detección de secuencias codificantes

Si bien la identificación de marcos de lectura abiertos dentro de una secuencia de ADN es sencilla, la identificación de secuencias codificantes no lo es, ya que la célula traduce solo un subconjunto de todos los marcos de lectura abiertos a proteínas. [ 26 ] Actualmente, la predicción de CDS utiliza el muestreo y la secuenciación de ARNm de las células, aunque aún existe el problema de determinar qué partes de un ARNm dado se traducen realmente a proteínas. La predicción de CDS es un subconjunto de la predicción de genes , que también incluye la predicción de secuencias de ADN que codifican no solo para proteínas, sino también para otros elementos funcionales como genes de ARN y secuencias reguladoras.
Tanto en procariotas como en eucariotas , la superposición de genes ocurre con relativa frecuencia en virus de ADN y ARN como una ventaja evolutiva para reducir el tamaño del genoma manteniendo la capacidad de producir diversas proteínas a partir de las regiones codificantes disponibles. [ 27 ] [ 28 ] Para ambos, los alineamientos por pares pueden detectar regiones codificantes superpuestas, incluyendo marcos de lectura abiertos cortos en virus, pero requerirían una cadena codificante conocida para comparar la posible cadena codificante superpuesta. [ 29 ] Un método alternativo que utiliza secuencias de genoma único no requeriría múltiples secuencias de genoma para realizar comparaciones, pero requeriría al menos 50 nucleótidos superpuestos para ser sensible. [ 30 ]
Véase también
- Cadena codificante: La cadena de ADN que codifica una proteína.
- Exón: La porción completa de la cadena que se transcribe.
- ARNm maduro: La porción del producto de transcripción del ARNm que se traduce.
- Estructura genética Los demás elementos que componen un gen
- Gen anidado: La secuencia codificante completa se encuentra dentro de los límites de un gen externo más grande.
- ADN no codificante Partes de los genomas que no codifican genes que codifican proteínas.
- Moléculas de ARN no codificante que no codifican proteínas, por lo que no tienen CDS.
- ADN no funcional Partes de los genomas sin función biológica relevante
Referencias
- 1 2 Twyman, Richard (1 de agosto de 2003). "Estructura genética" . The Wellcome Trust. Archivado del original el 28 de marzo de 2007. Recuperado el 6 de abril de 2003 .
- ↑ Höglund M, Säll T, Röhme D (febrero de 1990). "Sobre el origen de las secuencias codificantes a partir de marcos de lectura abiertos aleatorios". Journal of Molecular Evolution . 30 (2): 104– 108. Bibcode : 1990JMolE..30..104H . doi : 10.1007/bf02099936 . ISSN 0022-2844 . S2CID 5978109 .
- ↑ Sakharkar MK, Chow VT, Kangueane P (2004). "Distribuciones de exones e intrones en el genoma humano". In Silico Biology . 4 (4): 387– 93. doi : 10.3233/ISB-00142 . PMID 15217358 .
- ↑ Parnell, Laurence D. (2012-01-01). "Avances en tecnologías y diseño de estudios" . En Bouchard, C.; Ordovas, JM (eds.). Avances recientes en nutrigenética y nutrigenómica . Vol. 108. Academic Press. pp. 17–50 . doi : 10.1016/B978-0-12-398397-8.00002-2 . ISBN 9780123983978. PMID 22656372 . Consultado el 07-11-2019 .
{{cite book}}:|journal=ignorado ( ayuda ) - ↑ Gilbert W (febrero de 1978). "¿Por qué los genes están en pedazos?" . Nature . 271 (5645): 501. Bibcode : 1978Natur.271..501G . doi : 10.1038/271501a0 . PMID 622185 . S2CID 4216649 .
- ↑ Lercher MJ, Urrutia AO, Pavlícek A, Hurst LD (octubre de 2003). "Una unificación de estructuras mosaico en el genoma humano" . Human Molecular Genetics . 12 (19): 2411– 5. doi : 10.1093/hmg/ddg251 . PMID 12915446 .
- ↑ Oliver JL, Marín A (septiembre de 1996). "Una relación entre el contenido de GC y la longitud de la secuencia codificante". Journal of Molecular Evolution . 43 (3): 216– 23. Bibcode : 1996JMolE..43..216O . doi : 10.1007/pl00006080 . PMID 8703087 .
- ↑ "ROSALIND | Glosario | Región codificante del gen" . rosalind.info . Consultado el 31 de octubre de 2019 .
- ↑ Vinogradov AE (abril de 2003). "Hélice de ADN: la importancia de ser rica en GC" . Nucleic Acids Research . 31 (7): 1838– 44. doi : 10.1093/nar/gkg296 . PMC 152811. PMID 12654999 .
- ↑ Bohlin J, Eldholm V, Pettersson JH, Brynildsrud O, Snipen L (febrero de 2017). "La composición de nucleótidos de los genomas microbianos indica patrones diferenciales de selección en los genomas centrales y accesorios" . BMC Genomics . 18 (1) 151. doi : 10.1186 /s12864-017-3543-7 . PMC 5303225. PMID 28187704 .
- ↑ Sémon M, Mouchiroud D, Duret L (febrero de 2005). "Relación entre la expresión génica y el contenido de GC en mamíferos: significación estadística y relevancia biológica" . Human Molecular Genetics . 14 (3): 421–7 . doi : 10.1093/hmg/ddi038 . PMID 15590696 .
- 1 2 Descripción general de la transcripción. (s.f.). Recuperado de https://www.khanacademy.org/science/biology/gene-expression-central-dogma/transcription-of-dna-into-rna/a/overview-of-transcription .
- ↑ Clancy, Suzanne (2008). "Traducción: ADN a ARNm a proteína" . Scitable: Por Nature Education .
- ↑ Plociam (2005-08-08), Inglés: La estructura de un ARNm eucariota maduro. Un ARNm completamente procesado incluye la caperuza 5', la región 5' UTR, la región codificante, la región 3' UTR y la cola de poli(A). , consultado el 19-11-2019
- ↑ Shinohara K, Sasaki S, Minoshima M, Bando T, Sugiyama H (2006-02-13). "La alquilación de la cadena molde de la región codificante provoca un silenciamiento génico efectivo" . Nucleic Acids Research . 34 (4): 1189– 95. doi : 10.1093 / nar/gkl005 . PMC 1383623. PMID 16500890 .
- ↑ "Alquilación de ADN Término de ontología genética (GO:0006305)" . www.informatics.jax.org . Consultado el 30 de octubre de 2019 .
- ↑ Shafee T, Lowe R (2017). "Estructura genética eucariota y procariota" . WikiJournal of Medicine . 4 (1). doi : 10.15347/wjm/2017.002 .
- ↑ Konarska MM (1998). "Reconocimiento del sitio de empalme 5 'por parte del espliceosoma" . Acta Biochimica Polonica . 45 (4): 869– 81. doi : 10.18388/abp.1998_4346 . PMID 10397335 .
- ↑ Jonsta247 (10-05-2013), Inglés: Ejemplo de mutación silenciosa , consultado el 19-11-2019
{{citation}}: CS1 maint: nombres numéricos: lista de autores ( enlace ) - 1 2 Yang, J. (2016, 23 de marzo). ¿Qué son las mutaciones genéticas? Recuperado de https://www.singerinstruments.com/resource/what-are-genetic-mutation/ .
- 1 2 3 ¿Qué es una mutación genética y cómo ocurren las mutaciones? - Genetics Home Reference - NIH. (s.f.). Recuperado de https://medlineplus.gov/genetics/understanding/mutationsanddisorders/genemutation/ .
- ↑ "Corrección y reparación del ADN (artículo)" . Khan Academy . Consultado el 22 de mayo de 2023 .
- ↑ Peretó J. (2011) Hipótesis del bamboleo (Genética). En: Gargaud M. et al. (eds) Enciclopedia de Astrobiología. Springer, Berlín, Heidelberg
- 1 2 Havrilla, JM, Pedersen, BS, Layer, RM, & Quinlan, AR (2018). Un mapa de regiones codificantes restringidas en el genoma humano. Nature Genetics , 88–95. doi : 10.1101/220814
- ↑ Romiguier J, Roux C (2017). " Sesgos analíticos asociados con el contenido de GC en la evolución molecular" . Front Genet . 8 : 16. doi : 10.3389/fgene.2017.00016 . PMC 5309256. PMID 28261263 .
- ^ Furuno M, Kasukawa T, Saito R, Adachi J, Suzuki H, Baldarelli R, et al. (junio de 2003). "Anotación CDS en secuencia de ADNc completa" . Investigación del genoma . 13 (6B). Prensa del laboratorio de Cold Spring Harbor: 1478– 87. doi : 10.1101/gr.1060303 . PMC 403693 . PMID 12819146 .
- ↑ Rogozin IB, Spiridonov AN, Sorokin AV, Wolf YI, Jordan IK, Tatusov RL, Koonin EV (mayo de 2002). "Purificación y selección direccional en genes procariotas superpuestos" . Trends in Genetics . 18 (5): 228–32 . doi : 10.1016/S0168-9525(02)02649-5 . PMID 12047938 .
- ↑ Chirico N, Vianelli A, Belshaw R (diciembre de 2010). "Por qué se superponen los genes en los virus" . Actas. Ciencias Biológicas . 277 (1701): 3809–17 . doi : 10.1098/rspb.2010.1052 . PMC 2992710. PMID 20610432 .
- ↑ Firth AE, Brown CM (febrero de 2005). "Detección de secuencias codificantes superpuestas con alineamientos por pares" . Bioinformatics . 21 (3): 282– 92. doi : 10.1093/bioinformatics/bti007 . PMID 15347574 .
- ↑ Schlub TE, Buchmann JP, Holmes EC (octubre de 2018). Malik H (ed.). "Un método simple para detectar genes superpuestos candidatos en virus utilizando secuencias de genoma único" . Biología molecular y evolución . 35 (10): 2572–2581 . doi : 10.1093/molbev/msy155 . PMC 6188560. PMID 30099499 .
- ADN
- Bioquímica