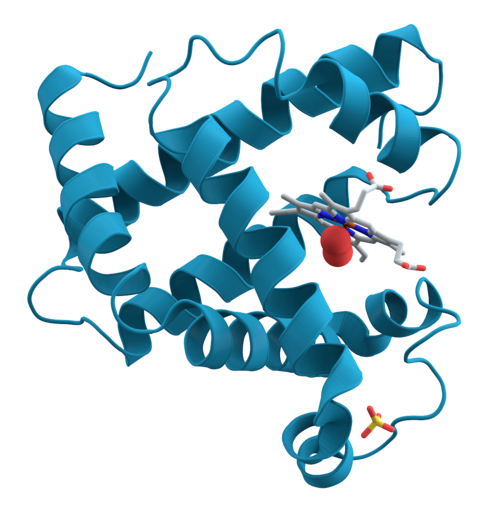Un péptido señal (a veces denominado secuencia señal , señal de direccionamiento , señal de localización , secuencia de localización , péptido de tránsito , secuencia líder o péptido líder ) es un péptido corto (generalmente de 16 a 30 aminoácidos de longitud) [ 1 ] presente en el extremo N-terminal (o, ocasionalmente, de forma no clásica, en el extremo C-terminal [ 2 ] o internamente) de la mayoría de las proteínas recién sintetizadas que están destinadas a la vía secretora . [ 3 ] Estas proteínas incluyen aquellas que residen dentro de ciertos orgánulos (el retículo endoplasmático , el aparato de Golgi o los endosomas ), secretadas por la célula o insertadas en la mayoría de las membranas celulares. Aunque la mayoría de las proteínas de membrana de tipo I tienen péptidos señal, la mayoría de las proteínas de membrana de tipo II y las proteínas de membrana multitransmembrana están dirigidas a la vía secretora por su primer dominio transmembrana , que bioquímicamente se asemeja a una secuencia señal excepto que no se escinde. Son un tipo de péptido diana .
Función (translocación)
Los péptidos señal funcionan para inducir a una célula a translocar la proteína, generalmente a la membrana celular. En procariotas , los péptidos señal dirigen la proteína recién sintetizada al canal conductor de proteínas SecYEG, que se encuentra en la membrana plasmática . Existe un sistema homólogo en eucariotas , donde el péptido señal dirige la proteína recién sintetizada al canal Sec61, que comparte homología estructural y de secuencia con SecYEG, pero se encuentra en el retículo endoplasmático. [ 4 ] Tanto el canal SecYEG como el Sec61 se conocen comúnmente como translocador , y el tránsito a través de este canal se denomina translocación. Mientras las proteínas secretadas se introducen a través del canal, los dominios transmembrana pueden difundirse a través de una compuerta lateral en el translocador para distribuirse en la membrana circundante.
Estructura
El núcleo del péptido señal contiene una larga secuencia de aminoácidos hidrofóbicos (de aproximadamente 5 a 16 residuos) [ 5 ] que tiende a formar una hélice alfa simple y también se denomina "región h". Además, muchos péptidos señal comienzan con una corta secuencia de aminoácidos con carga positiva, que puede ayudar a mantener la topología adecuada del polipéptido durante la translocación mediante la regla del "interior positivo" [ 6 ] . Debido a su proximidad al extremo N-terminal, se denomina "región n". Al final del péptido señal suele haber una secuencia de aminoácidos que es reconocida y escindida por la peptidasa señal , por lo que se denomina sitio de escisión. Este sitio de escisión está ausente en los dominios transmembrana que actúan como péptidos señal, a los que a veces se denomina secuencias de anclaje de señal. La peptidasa señal puede escindir durante o después de la translocación para generar un péptido señal libre y una proteína madura. Los péptidos señal libres son posteriormente digeridos por proteasas específicas. Además, distintos tipos de péptidos señal se dirigen a diferentes localizaciones. Por ejemplo, la estructura de un péptido dirigido al entorno mitocondrial difiere en longitud y presenta un patrón alternante de segmentos pequeños con carga positiva e hidrofóbicos. Los péptidos señal dirigidos al núcleo se encuentran tanto en el extremo N como en el extremo C de una proteína y, en la mayoría de los casos, se conservan en la proteína madura.
Translocación cotraduccional versus postraduccional
Tanto en procariotas como en eucariotas, las secuencias señal pueden actuar de forma cotraduccional o postraduccional.
La vía cotraduccional se inicia cuando el péptido señal emerge del ribosoma y es reconocido por la partícula de reconocimiento de señal (SRP). [ 7 ] La SRP detiene entonces la traducción (la detención de la traducción solo ocurre en eucariotas) y dirige el complejo secuencia señal-ribosoma-ARNm al receptor de SRP , que se encuentra en la superficie de la membrana plasmática (en procariotas) o del RE (en eucariotas). [ 8 ] Una vez completada la localización en la membrana, la secuencia señal se inserta en el translocón. Los ribosomas se acoplan físicamente a la cara citoplasmática del translocón y se reanuda la síntesis de proteínas. [ 9 ]
La vía postraduccional se inicia una vez completada la síntesis de proteínas. En procariotas, la secuencia señal de los sustratos postraduccionales es reconocida por la proteína chaperona SecB , que transfiere la proteína a la ATPasa SecA , la cual, a su vez, bombea la proteína a través del translocador. Si bien se sabe que la translocación postraduccional ocurre en eucariotas, aún no se comprende completamente. Se sabe que en levaduras, la translocación postraduccional requiere el translocador y dos proteínas adicionales unidas a la membrana: Sec62 y Sec63 . [ 10 ]
Determinación de la eficiencia de secreción
Los péptidos señal son extremadamente heterogéneos; muchos de los procariotas y eucariotas son funcionalmente intercambiables dentro o entre especies, y todos determinan la eficiencia de la secreción de proteínas. [ 11 ] [ 12 ] [ 13 ]
Características a nivel de nucleótidos
En los vertebrados, la región del ARNm que codifica el péptido señal (es decir, la región codificante de la secuencia señal, o SSCR) puede funcionar como un elemento de ARN con actividades específicas. Las SSCR promueven la exportación nuclear del ARNm y su correcta localización en la superficie del retículo endoplasmático. Además, las SSCR presentan características de secuencia específicas: tienen un bajo contenido de adenina , están enriquecidas en ciertos motivos y tienden a estar presentes en el primer exón con una frecuencia mayor de la esperada. [ 14 ] [ 15 ]
Mecanismos de secreción alternativos
Las proteínas sin péptidos señal también pueden secretarse mediante mecanismos no convencionales. Por ejemplo, la interleucina y la galectina. [ 16 ] El proceso por el cual estas proteínas secretoras acceden al exterior celular se denomina secreción proteica no convencional (UPS). En las plantas, hasta el 50 % de las proteínas secretadas pueden depender de la UPS. [ 17 ]
Secuencias no clásicas
Los péptidos señal suelen estar ubicados en el extremo N-terminal de las proteínas. Algunas presentan péptidos señal internos o en el extremo C-terminal (por ejemplo: señal de localización peroxisomal y señal de localización nuclear). La estructura de estos péptidos señal no clásicos difiere enormemente de la de los péptidos señal N-terminales. [ 2 ]
Nomenclatura
Los péptidos señal no deben confundirse con los péptidos líderes, a veces codificados por el ARNm líder, aunque ambos se denominan indistintamente "péptidos líderes". Estos otros péptidos líderes son polipéptidos cortos que no intervienen en la localización de proteínas, sino que regulan la transcripción o la traducción de la proteína principal, y no forman parte de la secuencia proteica final. Este tipo de péptido líder se refiere principalmente a una forma de regulación génica presente en bacterias, si bien se utiliza un mecanismo similar para regular genes eucariotas, denominado uORF (marcos de lectura abiertos corriente arriba).
El péptido señal como diana terapéutica
El péptido señal es una diana antiviral potencial (terapéutica) . Los péptidos señal estables (SSP) de los precursores de la glicoproteína viral del mammarenavirus se reasocian con las proteínas de la espícula maduras después de su escisión cotraduccional. [ 18 ] Los SSP con una glicina en el extremo N penúltimo son una diana para los inhibidores de NMT , [ 19 ] que inhiben la miristoilación de los péptidos señal y dirigen el péptido señal a la degradación proteasómica , afectando la fusión virus-célula . [ 18 ]
Véase también
Referencias
- ↑ Kapp, Katja; Schrempf, Sabrina; Lemberg, Marius K.; Dobberstein, Bernhard (1 de enero de 2013). Funciones posteriores a la focalización de los péptidos señal . Biociencia de las Landas.
- 1 2 Owji, Hajar; Nezafat, Navid; Negahdaripour, Manica; Hajiebrahimi, Ali; Ghasemi, Younes (agosto de 2018). "Una revisión exhaustiva de los péptidos señal: estructura, funciones y aplicaciones". European Journal of Cell Biology . 97 (6): 422– 441. doi : 10.1016/j.ejcb.2018.06.003 . PMID 29958716. S2CID 49612506 .
- ↑ Blobel G, Dobberstein B (diciembre de 1975). "Transferencia de proteínas a través de membranas. I. Presencia de cadenas ligeras de inmunoglobulina nacientes procesadas proteolíticamente y no procesadas en ribosomas unidos a la membrana de mieloma murino" . The Journal of Cell Biology . 67 (3): 835– 51. doi : 10.1083/jcb.67.3.835 . PMC 2111658. PMID 811671 .
- ↑ Rapoport TA (noviembre de 2007). "Translocación de proteínas a través del retículo endoplasmático eucariota y las membranas plasmáticas bacterianas". Nature . 450 ( 7170): 663– 9. Bibcode : 2007Natur.450..663R . doi : 10.1038/nature06384 . PMID 18046402. S2CID 2497138 .
- ↑ Käll L, Krogh A, Sonnhammer EL (mayo de 2004). "Un método combinado de predicción de topología transmembrana y péptido señal". Journal of Molecular Biology . 338 (5): 1027– 36. doi : 10.1016/j.jmb.2004.03.016 . PMID 15111065 .
- ↑ von Heijne G , Gavel Y (julio de 1988). "Señales topogénicas en proteínas de membrana integrales" . European Journal of Biochemistry . 174 (4): 671– 8. doi : 10.1111/j.1432-1033.1988.tb14150.x . PMID 3134198 .
- ↑ Walter P, Ibrahimi I, Blobel G (noviembre de 1981). "Translocación de proteínas a través del retículo endoplasmático. I. La proteína de reconocimiento de señal (SRP) se une a polisomas ensamblados in vitro que sintetizan proteína secretora" . The Journal of Cell Biology . 91 (2 Pt 1): 545–50 . doi : 10.1083/jcb.91.2.545 . PMC 2111968. PMID 7309795 .
- ↑ Gilmore R, Blobel G, Walter P (noviembre de 1982). "Translocación de proteínas a través del retículo endoplasmático. I. Detección en la membrana microsómica de un receptor para la partícula de reconocimiento de señal" . The Journal of Cell Biology . 95 (2 Pt 1): 463–9 . doi : 10.1083/jcb.95.2.463 . PMC 2112970. PMID 6292235 .
- ↑ Görlich D, Prehn S, Hartmann E, Kalies KU, Rapoport TA (octubre de 1992). "Un homólogo de mamíferos de SEC61p y SECYp se asocia con ribosomas y polipéptidos nacientes durante la translocación". Cell . 71 (3): 489– 503. doi : 10.1016/0092-8674(92)90517- G . PMID 1423609. S2CID 19078317 .
- ↑ Panzner S, Dreier L, Hartmann E, Kostka S, Rapoport TA (mayo de 1995). "Transporte de proteínas postraduccional en levadura reconstituida con un complejo purificado de proteínas Sec y Kar2p" . Cell . 81 ( 4): 561–70 . doi : 10.1016/0092-8674(95)90077-2 . PMID 7758110. S2CID 14398668 .
- ↑ Kober L, Zehe C, Bode J (abril de 2013). "Péptidos señal optimizados para el desarrollo de líneas celulares CHO de alta expresión". Biotecnología y bioingeniería . 110 (4): 1164– 73. Bibcode : 2013BiotB.110.1164K . doi : 10.1002/bit.24776 . PMID 23124363. S2CID 449870 .
- ↑ von Heijne G (julio de 1985). "Secuencias de señales. Los límites de la variación". Journal of Molecular Biology . 184 (1): 99– 105. doi : 10.1016/0022-2836(85)90046-4 . PMID 4032478 .
- ↑ Molino JV, de Carvalho JC, Mayfield SP (2018-02-06). "Comparación de péptidos señal secretores para la expresión de proteínas heterólogas en microalgas: Ampliando el portafolio de secreción para Chlamydomonas reinhardtii" . PLOS ONE . 13 (2) e0192433. Bibcode : 2018PLoSO..1392433M . doi : 10.1371/journal.pone.0192433 . PMC 5800701. PMID 29408937 .
- ↑ Palazzo AF, Springer M, Shibata Y, Lee CS, Dias AP, Rapoport TA (diciembre de 2007). "La región codificante de la secuencia señal promueve la exportación nuclear del ARNm" . PLOS Biology . 5 (12) e322. doi : 10.1371/journal.pbio.0050322 . PMC 2100149. PMID 18052610 .
- ↑ Cenik C, Chua HN, Zhang H, Tarnawsky SP, Akef A, Derti A, et al. (abril de 2011). Snyder M (ed.). "El análisis del genoma revela la interacción entre los intrones 5'UTR y la exportación nuclear de ARNm para genes secretorios y mitocondriales" . PLOS Genetics . 7 (4) e1001366. doi : 10.1371/journal.pgen.1001366 . PMC 3077370. PMID 21533221 .
- ↑ Nickel W, Seedorf M (2008). "Mecanismos no convencionales de transporte de proteínas a la superficie celular de células eucariotas". Annual Review of Cell and Developmental Biology . 24 : 287–308 . doi : 10.1146/annurev.cellbio.24.110707.175320 . PMID 18590485 .
- ↑ Agrawal GK, Jwa NS, Lebrun MH, Job D, Rakwal R (febrero de 2010). "Secretoma vegetal: desvelando los secretos de las proteínas secretadas". Proteomics . 10 ( 4): 799– 827. doi : 10.1002/pmic.200900514 . PMID 19953550. S2CID 20647387 .
- ^ Ingenio , Haydar; Betancourt, Carlos Alberto; Cubitt, Beatriz; Khafaji, Roaa; Kowalski, Heinrich; Jackson, Nathaniel; Sí, Chengjin; Martínez-Sobrido, Luis; de la Torre, Juan C. (2024-08-26). "Se requieren N-miristoiltransferasas celulares para la multiplicación del Mammarenavirus" . Virus . 16 (9): 1362. doi : 10.3390/v16091362 . ISSN 1999-4915 . PMC 11436053 . PMID 39339839 .
- ↑ Timms, Richard T.; Zhang, Zhiqian; Rhee, David Y.; Harper, J. Wade; Koren, Itay; Elledge, Stephen J. (2019). "Una vía de N-degrón específica de glicina media el control de calidad de la N- miristoilación de proteínas" . Science . 365 ( 6448 ) eaaw4912. doi : 10.1126/science.aaw4912 . PMC 7090375. PMID 31273098 .
Enlaces externos
- Péptido señal en los encabezamientos de materias médicas (MeSH) de la Biblioteca Nacional de Medicina de EE. UU.
- SPdb (Base de datos de péptidos señal) Archivada el 22/01/2016 en Wayback Machine
- SignalP predice la presencia y la ubicación de los sitios de escisión del péptido señal en secuencias de aminoácidos de diferentes organismos.
- expresión génica
- Orientación de proteínas